
It’s been a bumper summer for hihi research! I had two papers come out in the last couple of months, in which I explored the effects of a carotenoid supplementation experiment on a couple of different aspects of hihi breeding biology. Carotenoids are antioxidant pigments that birds must obtain through their diet. They are important in animal colouration, and are thought to play a role in immune health too (1, 2). The Tiritiri Matangi hihi population was supplemented with carotenoids supplied to the birds in sugar water feeders for two breeding seasons (2005-2007) in order to test whether mothers and/or offspring having access to more of these nutrients influenced their eggs, health, and provisioning behaviour. I was interested to follow up on these results by testing how carotenoid supplementation influenced:
- brood hierarchies
- and offspring sex ratio
First, what are brood hierarchies?

A brood hierarchy refers to size differences between chicks in a nest (brood = chicks in a nest). Brood hierarchies can arise if eggs differed in size or contents, but most often are thought to arise due to differences in when the eggs hatched, with eggs hatching first resulting in chicks that get a headstart and end up bigger than later-hatched siblings. This usually means they are more competitive, for example, being more successful in obtaining food from their parents.
Mothers can control how long their clutch (clutch = eggs in a nest) takes to hatch by adjusting when they begin sitting on their eggs to begin incubation. Birds lay one egg per day. If they begin incubation as soon as the first egg is laid, as below, the first egg will have a head start over those laid later as it will receive more incubation time and so will develop faster. This will usually mean that the first egg will hatch sooner, resulting in a brood hierachy:

If females wait to begin incubating their clutch until after the last egg is laid, as shown here, then all the eggs will develop at the same rate, and are likely to hatch at the same time:

(Another way to think about this – imagine you are boiling 3 eggs. If you put them all into boiling water at the same time, they’ll all be ready at the same time. If you put them in sequentially, a minute apart, the one you put in first will be hard-boiled before the others!)
Why are brood hierarchies important, and what can we do about them?
As strange as it seems, given that generating size differences within a brood is likely to result in later-hatched chicks being less competitive and perhaps less likely to survive, brood hierarchies are thought to be evolutionarily adaptive. For example, in uncertain environments, encouraging sibling competition within the nest may mean that at least the older, more competitive chicks are likely to survive and thrive – as opposed to having a nest of equally poor quality chicks that may all die. In a species of critical conservation concern like the hihi, however, brood hierarchies are not what we want to see, because every chick is vitally important!
I was interested to see if supplementing mothers with carotenoids while they were laying their eggs might mitigate the negative effects of hatching later. Later hatched chicks are often smaller, lighter, and less likely to survive – perhaps increased maternal access to carotenoids, a nutrient we know boosts chick health (3), might lessen some of these effects?
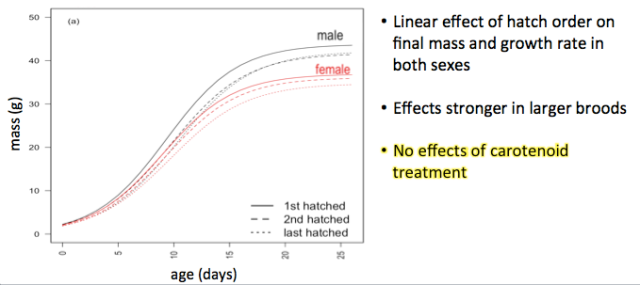
In our recent paper in Animal Behaviour, we show that carotenoid supplementation had no influence on the effects of brood hierarchy position in hihi. We did find strong hatch order effects – later hatched chicks were smaller, lighter, and less likely to survive as predicted (see figure below). This was an interesting result, as we found that the overall degree of hatch spread was very low relative to other species – usually all eggs hatched within 24 hours of each other, meaning that hihi are technically a “synchronously” hatching species! Perhaps we have previously underestimated hatch order effects in species where chicks all hatch relatively synchronously?
Carotenoids and offspring sex ratios – different benefits to males and females
Hihi are sexually dimorphic – males have distinctive yellow wing bars and white “eyebrow tufts”, both of which play a role in mate attraction and territory acquisition (4). Previous work has shown that dietary carotenoids are important in male yellow plumage (5), the colour of which influences the likelihood of being cuckolded (4). In other words – the availability of carotenoids in a male hihi’s early life can have big effects on his reproductive success as an adult. So, if mothers have a lot of carotenoids in their diet, in theory they should benefit most from producing lots of sons that will go on to have many offspring and not be cuckolded. So, does this actually happen?

Unfortunately, it’s hard to accurately test this question in many systems because often chicks die in the nest before researchers can identify their sexes, so commonly we know the sex ratio of a brood when they are ready to leave the nest, but not the sex ratio that was actually laid. Luckily, I was able to use tissue samples from chicks that died in the nest to obtain full sex ratios for each brood. You can read more about this lab work and what it entailed here!
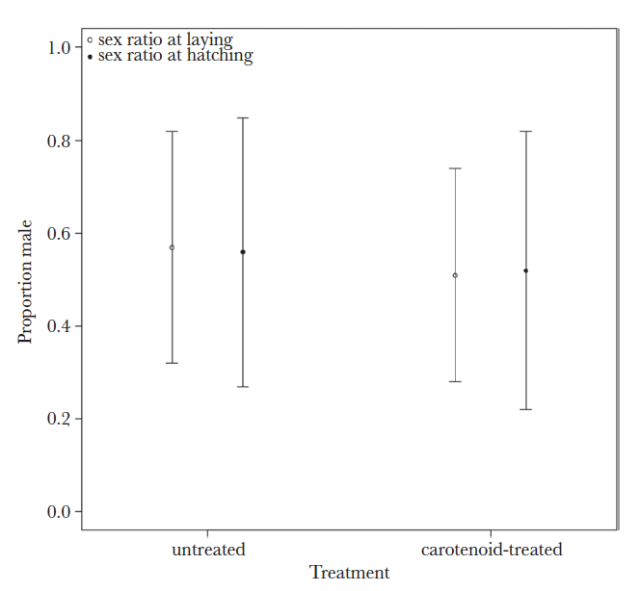
We found (published recently in Behavioral Ecology) that despite our predictions that, because males apparently benefit more from having more carotenoids in early life in terms of their long-term reproductive success, supplementing mothers with carotenoids did not alter offspring sex ratios (see graph below). Although this did not match our hypothesis, this is an important result – carotenoids have been shown to influence offspring sex ratios in other species (e.g. 6), and it’s equally important to report where this result is not replicated so we can better understand the role of carotenoids.
Why might this be? A few possibilities: perhaps carotenoids are actually also important to female long-term success in ways that we don’t know about yet, meaning that it’s less beneficial for supplemented mothers to invest more in males. For example, if the yellow colouration of their beaks is important in competition with other females, as is the case in goldfinches (7). It’s also possible that our carotenoid supplementation wasn’t a sufficient cue of carotenoid availability in the environment – not enough to have the big effects on male colouration that would result in important differences. Plenty to think about for the future!
Thanks to hihi collaborators!
On a personal note, it’s wonderful to see these hihi papers out, as they were great fun to work on, and the result of a great collaboration between Cambridge and John and Patricia at ZSL. Our funders for these projects are listed below.
As always, please do let me know if you would like to read any of the papers I’ve talked about and you are unable to access them. You can find my email in my contact details tab above.

Key resources:
Hihi conservation – updates, news, and information
The Informed Birds research group
References:
- Blount JD. 2004. Carotenoids and life-history evolution in animals. Arch Biochem Biophys. 430:10–15.
- Svensson PA, Wong BBM. 2011. Carotenoid-based signals in behavioural ecology: a review. Behaviour. 148:131–189.
- Ewen JG, Thorogood R, Brekke P, Cassey P, Karadas F, Armstrong DP. 2009. Maternally invested carotenoids compensate costly ectoparasitism in the hihi. Proc Natl Acad Sci USA. 106:12798–12802.
- Walker LK, Thorogood R, Karadas F, Raubenheimer D, Kilner RM, Ewen JG. 2014. Foraging for carotenoids: do colorful male hihi target carotenoid-rich foods in the wild? Behav Ecol. 25:1048–1057.
- Walker LK, Stevens M, Karadaş F, Kilner RM, Ewen JG. 2013. A window on the past: male ornamental plumage reveals the quality of their early life environment. Proc Roy Soc Lond B Biol Sci. 280:20122852.
- McGraw KJ, Adkins-Regan E, Parker RS. 2005. Maternally derived carotenoid pigments affect offspring survival, sex ratio, and sexual attractiveness in a colorful songbird. Die Naturwissenschaften. 92:375–380.
- Murphy TG, Rosenthal MF, Montgomerie R, Tarvin K. 2009. Female American goldfinches use carotenoid-based bill coloration to signal status.Behav Ecol. 20:1348–1355.
